
Comparative Pathology of Nonhuman Primate Atherosclerosis
A Historical Critical Review of Progression, Stabilization, Regression, and Plaque-Lipid Dynamics (1950–1995)
Executive Summary
The paradigm of atherosclerosis in the mid-twentieth century transitioned from viewing the condition as an inevitable, progressive consequence of aging to understanding it as a dynamic, lipid-driven inflammatory disease in which substantial regression and stabilization are biologically possible under some experimental conditions. Much of the foundational evidence for this shift was established through nonhuman primate models developed between 1950 and the late 1980s. Unlike lower mammalian models such as rabbits and rodents, nonhuman primates possess lipoprotein distributions, apolipoprotein B (ApoB) biology, and coronary arterial structures that closely resemble those of humans. This historically grounded scientific review examines the progression, stabilization, and regression of diet-induced atherosclerosis across several primate species, including rhesus macaques, cynomolgus macaques, African green monkeys, baboons, and squirrel monkeys.
The primary findings of these historical investigations demonstrate that nonhuman primates develop complex, human-like coronary plaques featuring fibrous caps, lipid-rich necrotic cores, extracellular matrix remodeling, calcification, and, less commonly, spontaneous myocardial infarction. Furthermore, serial and postmortem analyses provide compelling experimental evidence that reversing severe hypercholesterolemia—either through dietary lipid restriction or pharmacological and natural compounds—can induce significant plaque regression. This regression is characterized by the preferential and sometimes relatively rapid depletion of intracellular and extracellular cholesteryl ester pools, the resolution of necrotic debris, and the restoration of endothelial vasomotor function, although densely fibrotic and macrocalcific matrix components frequently persist, limiting complete anatomical normalization. By evaluating the dietary, kinetic, and methodological parameters of these historical studies, this report provides a translational bridge to modern human trials, outlining both the clinical potential and the biological limits of regression therapy.
Historical Evolution and Timeline
The systematic study of nonhuman primate atherosclerosis evolved through distinct experimental phases, tracking advancements in analytical biochemistry, histopathology, and imaging technologies. The table below outlines the milestones of this research, detailing how investigators advanced from early observation of vascular lipid deposits to functional, high-resolution evaluations of plaque biology.
| Period | Experimental Focus & Methodological Advancements | Key Pioneering Research Centers | Landmark Scientific Contributions |
| 1950s | Induction of hypercholesterolemia and vascular lipid staining in New World primates; exploration of nutritional deficiencies. | Harvard School of Public Health; Bowman Gray School of Medicine. | First induction of hypercholesterolemia in New World monkeys; characterization of Cebus and rhesus vascular susceptibility. |
| 1960s | Focus on coronary artery pathology and natural history; early reports of spontaneous myocardial infarction in hyperlipidemic monkeys. | Northwestern University; Oregon Regional Primate Research Center. | Taylor et al. document fatal diet-induced myocardial infarction in a rhesus macaque [1]. Maruffo & Portman study early coronary changes in squirrel monkeys [2]. |
| 1970s | Definitive morphometric proof of advanced coronary regression; detailed mapping of plaque lipid phases and matrix changes. | University of Iowa; University of Chicago; LSU Medical Center. | Armstrong et al. prove advanced coronary atheroma regression in rhesus macaques [3]. Vesselinovitch & Wissler evaluate low-fat diets combined with a pharmacologic lipid-lowering agent (W-1372) [4]. |
| 1980s | Long-term, multi-year kinetic studies of regression; physical-chemical analyses of lipid phase transitions; epidemiological modeling of HDL. | Bowman Gray School of Medicine; Boston University; Southwest Foundation for Biomedical Research. | Clarkson et al. define target cholesterol levels (200 vs. 300 mg/dl) for long-term regression [5]. Small et al. characterize transient plaque crystallization at 6 months of regression [6]. |
| 1990s | Integration of histomorphometry with quantitative angiography, endothelial reactivity, and compensatory vascular remodeling. | Bowman Gray School of Medicine; LSU Medical Center. | Williams et al. demonstrate that lipid lowering doubles coronary lumen area via outward remodeling and restores vasomotor response [7]. LSU team completes a multi-year study on rhesus plaques [8]. |
Primate Models: Species-Specific Pathobiology and Metabolic Profiles
The selection of a nonhuman primate species in atherosclerosis research is guided by genetic, metabolic, and anatomical considerations. Choosing the correct model is critical, as species differences in lipoprotein transport and arterial susceptibility directly affect the translation of experimental results to human clinical practice.
Rhesus Macaques (Macaca mulatta)
Rhesus macaques are historically the most widely utilized primate model for atherosclerosis research. Under dietary lipid challenge, they transition from an HDL-dominant state to an LDL-dominant state, with a plasma lipoprotein profile that closely mimics human hyperlipoproteinemia. Rhesus macaques are highly responsive to dietary cholesterol and saturated fat. They reliably develop advanced, concentric, and eccentric coronary plaques that progress to necrotic core formation, cellular decay, intraplaque hemorrhage, and calcification. Spontaneous myocardial infarction has been documented, though it remained uncommon, in long-term rhesus studies, making this species a leading model for clinical coronary artery disease.
Cynomolgus Macaques (Macaca fascicularis)
Cynomolgus macaques are highly sensitive to dietary cholesterol, making them an excellent model for rapid lesion induction. They are prone to developing advanced coronary arterial lesions with a high proportion of connective tissue, including collagen, elastin, and glycosaminoglycans, as well as dense calcification. Cynomolgus monkeys also exhibit a high degree of psychosocial stress reactivity. In dominant males and socially subordinate females, sympathetic nervous system activation and hypercortisolemia accelerate and worsen coronary artery atherogenesis, independent of circulating lipid levels. This unique neuroendocrine interaction makes them a valuable model for examining the relationship between lifestyle, stress, and vascular disease.
African Green Monkeys (Chlorocebus aethiops)
African green monkeys (vervets) possess a lipoprotein metabolism that is highly sensitive to the specific fatty acid composition of the diet. When fed monounsaturated versus saturated or polyunsaturated fats, they show clear variations in LDL size and density that mirror human dietary responses. Their coronary lesions develop more slowly than those of macaques, but they closely resemble human plaques in their distribution and their balanced cellular ratio of smooth muscle cells to macrophage foam cells.
Baboons (Papio hamadryas/sp.)
Baboons are large, genetically heterogeneous Old World primates widely used to study the genetic and environmental determinants of cardiovascular risk. They naturally develop early-stage fatty streaks and fibrous plaques at predictable arterial branches, including the iliac bifurcation and the abdominal aorta. Their growth, development, and aging follow a generalized Old World primate pattern, and they exhibit human-like variations in lipoprotein responses, blood pressure, and vascular compliance under dietary challenge. However, they are highly resistant to developing severe, occlusive coronary artery stenosis or terminal ischemic events within standard experimental timeframes, limiting their utility for advanced regression trials.
Squirrel Monkeys (Saimiri sciureus)
As small New World primates, squirrel monkeys offer a compact model for atherosclerosis research. They naturally develop aortic and coronary lesions, and are highly responsive to short-term dietary lipid manipulations. Reversion to low-cholesterol diets in squirrel monkeys induces rapid clearance of intracellular foam cells and early plaque regression. However, their small vessel caliber and overall physical dimension constraints complicate serial angiography, intravascular imaging, and regional hemodynamic flow measurements.
Comparative Advantages of Primate Models over Non-Primate Models
Nonhuman primates offer several physiological and structural advantages over lower mammalian models for questions about human coronary disease, although no single species is optimal for every purpose and other models (rabbits, pigs, and genetically modified mice) may be preferable for particular mechanistic, interventional, or imaging questions:
- Lipoprotein Distribution & CETP Activity: Rodents transport the vast majority of their circulating cholesterol in HDL particles and lack cholesteryl ester transfer protein (CETP) activity, which prevents them from developing human-like hyperlipidemia without massive genetic disruption. Many nonhuman-primate species possess CETP activity and, when challenged with dietary cholesterol and saturated fat, develop marked increases in ApoB-containing lipoproteins and a lipoprotein distribution resembling human hypercholesterolemia. Baseline lipoprotein distributions nevertheless vary substantially by species, colony, diet, sex, and individual responsiveness, and several primates are relatively HDL-dominant before cholesterol feeding.
- Arterial Anatomy and Hemodynamics: Nonhuman primates possess a multi-layered, thick coronary intima with a well-defined internal elastic lamina and adventitial vasa vasorum, matching human vascular architecture. Rabbits, by contrast, are extremely responsive to dietary cholesterol and readily accumulate lipid-laden macrophage foam cells (the “Anitschkow cell” response); under prolonged or modified protocols they can also develop more advanced fibrous, necrotic, and calcific lesions. Their extreme dietary-cholesterol sensitivity, distinctive lipoprotein metabolism, and frequent concentration of disease in the aorta rather than in spontaneous human-like coronary events nevertheless limit direct translation relative to selected primate models.
- Inflammatory and Thrombotic Cascades: Primates exhibit a highly conserved inflammatory cascade, utilizing closely homologous cell adhesion molecules (VCAM-1, ICAM-1), monocyte chemoattractants (MCP-1), and scavenger receptors (CD36, SR-A1) to drive macrophage recruitment and activation within the plaque. Furthermore, primate coagulation systems, platelet reactivity, and fibrinolytic cascades are highly comparable to those of humans, allowing for a more accurate modeling of plaque rupture, intraplaque hemorrhage, and luminal thrombosis than is possible in rodent or avian models.
Relevance Ranking of Major Research Programs
The table below presents an author-generated relevance ranking (not the output of a systematic review) of ten informative nonhuman primate research programs based on their relevance to human coronary atherosclerosis. The set includes both regression studies and comparative progression studies (notably the Rudel and McGill programs), which are labeled accordingly. Programs are ranked according to species suitability, the complexity and location of the induced lesions (coronary vs. aortic), the duration of the experimental feeding periods, the presence of a controlled regression phase, the measurement of plasma lipoprotein subfractions, and the detail of the postmortem coronary histopathology.
| Rank | Research Program & Lead Investigators | Primary Species | Coronary Lesion Complexity | Study Design & Kinetic Duration | Scientific Justification for Ranking |
| 1 | Iowa Coronary Regression Program (Armstrong, Warner, & Connor) | Macaca mulatta | High; severe diffuse coronary stenosis, necrosis, lipid-laden plaques. | 17 months induction; 40 months regression. | Gold Standard: First study to demonstrate substantial anatomical regression and lipid depletion in the coronary arteries of a primate model, demonstrating substantial reduction of diet-induced coronary atheromatosis and improved lumen dimensions. |
| 2 | LSU Long-Term Regression Program (Strong, Bhattacharyya, Eggen, Newman, et al.) | Macaca mulatta | High; structured, mature fibromuscular coronary plaques with lipid cores. | 5.4 years induction; 1.9 or 3.7 years regression. | Temporal Accuracy: Closest replication of the mature, decades-old plaque architecture of human adults; showed that regression of advanced lesions becomes demonstrable only after years of sustained lipid lowering. |
| 3 | Bowman Gray Primate Center Series V (Clarkson, Bond, Bullock, McLaughlin, Sawyer) | Macaca mulatta | High; concentric and eccentric coronary lesions with necrosis and calcification. | 38 months induction; 24 or 48 months regression. | Target Calibration: Defined target lipid levels during intervention; established that a plasma total cholesterol ceiling near 200 mg/dl is required for consistent coronary regression. |
| 4 | Bowman Gray Female Regression Program (Williams, Anthony, Honoré, Register, Clarkson, et al.) | Macaca fascicularis | High; complex calcified plaques, loss of endothelium-dependent vasomotor reactivity. | 24 months induction; 30 months regression. | Functional Integration: Showed that lipid lowering can approximately double the coronary lumen area—consistent with favorable outward remodeling—and restore vasomotor function even when plaque size does not shrink. |
| 5 | Boston / Bowman Gray Lipid Phase Program (Small, Bond, Waugh, Prack, Sawyer) | Macaca fascicularis | Moderate-High; epicardial coronary plaques, prominent cellular necrosis. | 18–30 months induction; 6 or 12 months regression. | Biophysical Insight: Characterized the physical chemistry of lipids in the plaque; described a transient crystalline free-cholesterol phase during early rapid lipid mobilization. |
| 6 | Chicago Reversal Program (Vesselinovitch, Wissler, Hughes, Borensztajn) | Macaca mulatta | Moderate-High; severe aortic and proximal coronary plaque. | 18 months induction; 18 months regression. | Therapeutic Synergy: Proved that combining low-fat diets with an added pharmacologic lipid-lowering agent (W-1372) accelerates foam cell clearance and promotes endothelial repair. |
| 7 | Oregon Alfalfa Saponin Program (Malinow, McLaughlin, Naito, McNulty, et al.) | Macaca fascicularis | Moderate; widespread coronary foam cell infiltration and early plaques. | 6 months induction; 18 months regression. | Mechanistic Diversification: Showed that blocking intestinal cholesterol absorption with plant extracts (saponins) can regress plaque without requiring a complete shift to a low-fat diet. |
| 8 | Wake Forest / Bowman Gray Fatty Acid Program (Rudel, Parks, Sawyer) | Chlorocebus aethiops | Moderate-High; proximal coronary plaques rich in cholesteryl esters. | 5 years continuous induction (comparative fat study). | Lipoprotein Biology: Challenged the assumption that dietary fats are equivalent; showed that monounsaturated fat did not protect and was associated with large, cholesteryl-oleate-enriched, atherogenic LDL. |
| 9 | Southwest Foundation Baboon Program (McGill, McMahan, Kruski, Mott) | Papio sp. | Low-Moderate; aortic and iliac bifurcational fatty streaks and early plaques. | 26 months continuous atherogenic induction. | Epidemiological Validation: Confirmed the positive association of LDL and negative association of HDL with plaque coverage under controlled conditions, validating human models. |
| 10 | Oregon New World Program (Maruffo & Portman) | Saimiri sciureus | Low-Moderate; early proximal coronary foam cell lesions. | 3–12 months induction; 3–12 months control-diet follow-up. | Early Validation: One of the first programs to examine the fate of early coronary foam-cell lesions in a New World primate after dietary lipid normalization. |
Individual Structured Evidence Profiles
The following structured profiles present the scientific data and pathological findings for each of the ranked research programs. Bibliographic details have been verified against the primary literature; quantitative values drawn from the original reports are presented as published.
Rank 1: Iowa Coronary Regression Program (Armstrong, Warner, & Connor, 1970)
| Full citation | [3] Armstrong M. L., Warner E. D., Connor W. E. Regression of coronary atheromatosis in rhesus monkeys. Circ Res. 1970 Jul;27(1):59–67. |
| Year | 1970 |
| Primate species | Macaca mulatta (Rhesus macaque) |
| Number of animals | 40 |
| Sex and approximate age | Adult males, age unknown (wild-caught) |
| Duration of atherogenic feeding | 17 months |
| Duration of regression phase | 40 months |
| Experimental diet composition | Semipurified diet containing 41% of total calories from fat, 19% from protein, and 40% from carbohydrate. |
| Control / regression diet | Low-fat diet (4% calories as fat) OR corn-oil diet (40% calories as corn oil). Both were cholesterol-free. |
| Dietary cholesterol content | 1.2% by weight during induction; 0% (cholesterol-free) during regression. |
| Total-fat percentage | 41% (induction), 40% (corn-oil regression), 4% (low-fat regression). |
| Saturated-fat percentage | High saturated fat during induction (egg-yolk fat); low saturated fat during corn-oil regression. |
| Achieved lipids & lipoproteins | Induction: 711 ± 31 mg/dl (TC). Low-fat regression: 141 ± 7 mg/dl. Corn-oil regression: 140 ± 8 mg/dl. Electrophoresis showed complete reversal of beta-lipoproteinemia. |
| Arterial territories examined | Extramural coronary arteries (LAD, LCx, RCA), aorta, and peripheral arteries. |
| Plaque-assessment method | Histomorphometry of microscope-projected sections traced on paper, cut, and weighed. |
| Evidence of coronary disease | Severe, diffuse coronary atheromatosis with marked luminal narrowing across five evaluation sites. |
| Evidence of plaque progression | High-fat, high-cholesterol feeding caused extensive progression of lesions in 17 months. |
| Evidence of true regression | The average cross-sectional area of the arterial lumen was more than 80% greater in regression animals than in animals with baseline atherosclerosis, with no significant difference between the low-fat and corn-oil diets. |
| Evidence of plaque stabilization | Remaining plaques were flat, depleted of lipid, and replaced by dense fibrous tissue. |
| Calcification / thrombosis / death | No fatal infarctions reported in this specific cohort; minimal baseline calcification observed. |
| Main conclusions | Uncomplicated coronary atheromas regress substantially when plasma lipids are normalized by low-fat or corn-oil diets. |
| Major limitations | Small sample sizes, lack of baseline coronary biopsies in surviving animals, and lack of modern lipoprotein assays. |
| Relevance to human disease | Very High; demonstrated the anatomical feasibility of reducing coronary stenosis via dietary modification. |
Rank 2: LSU Long-Term Regression Program (Strong, Bhattacharyya, Eggen, Newman, et al., 1994)
| Full citation | [8] Strong J. P., Bhattacharyya A. K., Eggen D. A., et al. Long-term induction and regression of diet-induced atherosclerotic lesions in rhesus monkeys. I & II. Arterioscler Thromb. 1994;14(6):958–965; 14(12):2007–2016. |
| Year | 1994 |
| Primate species | Macaca mulatta (Rhesus macaque) |
| Number of animals | 45 |
| Sex and approximate age | Young adult males, 3–6 years of age at baseline |
| Duration of atherogenic feeding | 5.4 years |
| Duration of regression phase | 1.9 years (Group R4) or 3.7 years (Group R5) |
| Experimental diet composition | High-saturated-fat, high-cholesterol diet: 38% of calories from fat, 15% from protein, and 47% from carbohydrate. |
| Control / regression diet | Saturated-fat diet without cholesterol (38% fat, 15% protein, 47% carbohydrate; cholesterol deleted). |
| Dietary cholesterol content | 0.35 mg/kcal during induction; 0.02 mg/kcal during regression. |
| Total-fat percentage | 38.0% of total calories |
| Saturated-fat percentage | 26.2% of total calories (polyunsaturated/saturated ratio of 0.35). |
| Achieved lipids & lipoproteins | Serum cholesterol rose from a pre-induction mean near 150 mg/dl to a mean of about 430 mg/dl during induction, and returned toward baseline (approximately 150 mg/dl) during regression. |
| Arterial territories examined | Aorta, coronary arteries (LAD, LCx, RCA), common carotid, external carotid, and peripheral arteries. |
| Plaque-assessment method | Visual estimation of aortic surface lesion area, chemical lipid extraction, and histomorphometry of perfusion-fixed sections. |
| Evidence of coronary disease | Concentric and eccentric plaques with measurable coronary luminal reduction at baseline. |
| Evidence of plaque progression | 5.4 years of induction produced extensive progression of raised lesions and elevated plaque cholesteryl esters. |
| Evidence of true regression | Statistically demonstrable decrease in intimal thickness and plaque area became apparent after approximately 3.7 years of regression (Group R5), but not at 1.9 years. |
| Evidence of plaque stabilization | Substantial reduction in plaque esterified cholesterol, with remaining lesions composed of dense collagenous scars. |
| Calcification / thrombosis / death | Advanced calcification in the abdominal aorta and proximal coronaries, which remained largely unchanged during regression. |
| Main conclusions | Advanced, diet-induced coronary plaques can regress, but they require a significantly longer duration of lipid lowering than early-stage lesions. |
| Major limitations | Prolonged study length resulted in animal attrition; small sample size in late regression cohorts. |
| Relevance to human disease | Extremely High; closely models the chronicity, slow clearance, and structural maturity of human adult plaques. |
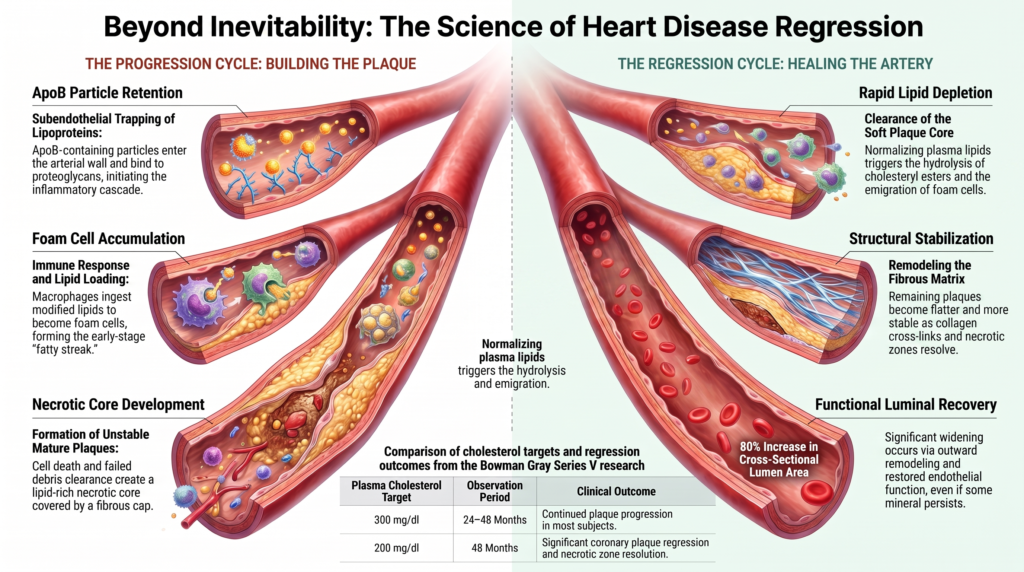
Rank 3: Bowman Gray Primate Center Series V (Clarkson, Bond, Bullock, McLaughlin, Sawyer, 1984)
| Full citation | [5] Clarkson T. B., Bond M. G., Bullock B. C., McLaughlin K. J., Sawyer J. K. A study of atherosclerosis regression in Macaca mulatta. V. Changes in abdominal aorta, carotid, and coronary arteries. Exp Mol Pathol. 1984 Aug;41(1):96–118. |
| Year | 1984 |
| Primate species | Macaca mulatta (Rhesus macaque) |
| Number of animals | 48 |
| Sex and approximate age | Young adult males, age approximately 4–5 years at baseline |
| Duration of atherogenic feeding | 38 months |
| Duration of regression phase | 24 or 48 months |
| Experimental diet composition | Semipurified atherogenic diet (approximately 40–44% of calories from fat, primarily lard). |
| Control / regression diet | Titrated semipurified diets where cholesterol content was adjusted individually to maintain specific plasma total-cholesterol ceilings. |
| Dietary cholesterol content | Approximately 0.35–1.0 mg/Cal during induction; dynamically titrated during regression. |
| Total-fat percentage | Approximately 40–44% (induction); lower fat during regression titrations. |
| Saturated-fat percentage | High saturated fat during induction (lard/beef tallow/butter). |
| Achieved lipids & lipoproteins | Induction: 450 ± 25 mg/dl (TC). Regression Group 1: 316 ± 10 mg/dl. Regression Group 2: 204 ± 4 mg/dl. |
| Arterial territories examined | Abdominal aorta, thoracic aorta, coronary arteries (LAD, LCx, RCA), and common carotid bifurcations. |
| Plaque-assessment method | Perfusion-fixed morphometry, detailed histological grading, and biochemical analysis of tissue cholesteryl esters. |
| Evidence of coronary disease | Concentric fibro-lipid coronary plaques with prominent necrotic cores and medial thinning. |
| Evidence of plaque progression | 38 months of induction produced severe coronary plaques. Progression continued in a substantial fraction of the 300 mg/dl cohort. |
| Evidence of true regression | Coronary plaque regressed significantly after 48 months at 200 mg/dl, but not after 24 months, showing time-dependence. |
| Evidence of plaque stabilization | Monkeys at 200 mg/dl showed near-complete resolution of necrotic zones and a relative increase in dense collagen matrix. |
| Calcification / thrombosis / death | High baseline calcification. Calcification frequency was substantially lower in the 200 mg/dl group than in the 300 mg/dl group. |
| Main conclusions | Reversing advanced coronary plaques requires maintaining a plasma total cholesterol ceiling near 200 mg/dl for several years. |
| Major limitations | High individual genetic variability (hyper- vs. hypo-responders) confounded findings in the 300 mg/dl regression cohort. |
| Relevance to human disease | High; established that absolute lipid targets and timeframes are critical factors for successful regression. |
Rank 4: Bowman Gray Female Regression Program (Williams, Anthony, Honoré, Register, Clarkson, et al., 1995)
| Full citation | [7] Williams J. K., Anthony M. S., Honoré E. K., Herrington D. M., Morgan T. M., Register T. C., Clarkson T. B. Regression of atherosclerosis in female monkeys. Arterioscler Thromb Vasc Biol. 1995 Jul;15(7):827–836. |
| Year | 1995 |
| Primate species | Macaca fascicularis (Cynomolgus macaque) |
| Number of animals | 88 (baseline necropsy n=20; diet-only n=25; +conjugated equine estrogens n=22; +estrogens/medroxyprogesterone n=21) |
| Sex and approximate age | Surgically postmenopausal (bilaterally ovariectomized) adult females |
| Duration of atherogenic feeding | 24 months |
| Duration of regression / treatment phase | 30 months |
| Experimental diet composition | Semipurified atherogenic diet (approximately 44% of calories from fat, 16% protein, 40% carbohydrate). |
| Control / regression diet | Low-fat, low-cholesterol diet enriched with safflower oil. |
| Dietary cholesterol content | High during induction; markedly reduced during the regression/treatment phase. |
| Achieved lipids & lipoproteins | Induction: approximately 470 mg/dl (TC). Regression: decreased to a mean near 140 mg/dl in the diet-only and hormone-added groups. |
| Arterial territories examined | Extramural coronary arteries (LAD, LCx, RCA) and abdominal aorta. |
| Plaque-assessment method | Perfusion-fixed coronary morphometry and vascular-ring vasomotor reactivity. |
| Evidence of coronary disease | Advanced, calcified epicardial coronary plaques causing baseline luminal narrowing. |
| Evidence of plaque progression | 24 months of induction produced severe concentric and eccentric coronary lesions. |
| Evidence of true regression | Cross-sectional plaque size did not change significantly compared with the baseline necropsy cohort. |
| Evidence of plaque stabilization | Coronary artery and lumen cross-sectional area approximately doubled compared with the baseline group, consistent with favorable (outward) remodeling; endothelium-dependent vasomotor dilation to acetylcholine was improved. |
| Calcification / thrombosis / death | Advanced calcification remained largely unchanged; its persistence limits anatomical normalization and does not by itself indicate healing. |
| Main conclusions | Clinically relevant benefit is not restricted to plaque shrinkage; lipid lowering was associated with enlargement of coronary artery and lumen cross-sectional area and improved endothelial reactivity, consistent with favorable remodeling. |
| Major limitations | Emphasis on postmenopausal hormone interactions; the advanced lesions did not show direct volumetric shrinkage. |
| Relevance to human disease | Extremely High; offers a plausible biological explanation for why lipid lowering reduces clinical events in humans despite minimal change in angiographic stenosis. |
Rank 5: Boston / Bowman Gray Lipid Phase Program (Small, Bond, Waugh, Prack, Sawyer, 1984)
| Full citation | [6] Small D. M., Bond M. G., Waugh D., Prack M., Sawyer J. K. Physicochemical and histological changes in the arterial wall of nonhuman primates during progression and regression of atherosclerosis. J Clin Invest. 1984 Jun;73(6):1590–1605. |
| Year | 1984 |
| Primate species | Macaca fascicularis (Cynomolgus macaque) |
| Number of animals | 54 |
| Sex and approximate age | Adult males, age approximately 5–6 years at baseline |
| Duration of atherogenic feeding | Up to 30 months (progression animals evaluated at 6, 12, 18, 24, and 30 months) |
| Duration of regression phase | 6 or 12 months (after 18 months of induction) |
| Experimental diet composition | Semipurified diet containing dextrin, salts, vitamin mixture, and butterfat/lard. |
| Control / regression diet | Standard commercial non-cholesterol-containing monkey chow. |
| Dietary cholesterol content | 0.36 mg/Cal during induction; cholesterol-free during regression. |
| Total-fat percentage | Approximately 40% of calories as fat during induction; low fat during chow regression. |
| Saturated-fat percentage | High (butterfat and lard) during induction. |
| Achieved lipids & lipoproteins | Induction: 520 ± 30 mg/dl (TC). 6-month regression: 148 ± 12 mg/dl. 12-month regression: 140 ± 10 mg/dl. |
| Arterial territories examined | Thoracic aorta, abdominal aorta, common carotid, and coronary arteries. |
| Plaque-assessment method | Perfusion histomorphometry, hot-stage polarizing microscopy, and chemical lipid extraction. |
| Evidence of coronary disease | Advanced, occlusive fibro-lipid epicardial coronary plaques with large necrotic cores. |
| Evidence of plaque progression | Plaque cholesteryl esters increased, forming foam-cell-rich lesions with high-melting-point intracellular lipid droplets. |
| Evidence of true regression | Return to chow shifted plaque lipid composition toward normal and reduced lesion size by 12 months. |
| Evidence of plaque stabilization | After 12 months of regression, cell debris and necrotic core size were significantly reduced, and collagen content increased. |
| Calcification / thrombosis / death | During early regression, a transient increase in free cholesterol monohydrate crystals was observed, interpreted as cholesteryl-ester hydrolysis temporarily exceeding cholesterol efflux. |
| Main conclusions | Plaque regression involves complex, time-dependent chemical changes; rapid lipid lowering can induce a transient crystalline phase during lipid mobilization. |
| Major limitations | Confined to a single species with rapid lipid-loading responses. |
| Relevance to human disease | High; elucidated the physical chemistry of plaque lipid mobilization, explaining why structural healing takes time to manifest. |
Rank 6: Chicago Reversal Program (Vesselinovitch, Wissler, Hughes, Borensztajn, 1976)
| Full citation | [4] Vesselinovitch D., Wissler R. W., Hughes R., Borensztajn J. Reversal of advanced atherosclerosis in rhesus monkeys. Part 1. Gross and light-microscopic studies. Atherosclerosis. 1976 Feb;23(2):155–176. |
| Year | 1976 |
| Primate species | Macaca mulatta (Rhesus macaque) |
| Number of animals | 54 |
| Sex and approximate age | Young adult males, 3–4 years of age at baseline |
| Duration of atherogenic feeding | 18 months |
| Duration of regression phase | 18 months |
| Experimental diet composition | Semipurified diet with 25% by weight of a 1:1 coconut-oil / butterfat mixture, plus casein and cornstarch. |
| Control / regression diet | Low-fat, low-cholesterol diet with or without the investigational lipid-lowering compound W-1372 (an experimental agent, not a clinically approved drug). |
| Dietary cholesterol content | 2.0% by weight during induction; cholesterol-free during regression. |
| Total-fat percentage | High saturated fat during induction; less than 5% during regression. |
| Saturated-fat percentage | High saturated fat during induction (coconut oil and butterfat). |
| Achieved lipids & lipoproteins | Serum cholesterol increased roughly five-fold during induction (approximately 830 ± 45 mg/dl). Regression (diet only): 210 ± 15 mg/dl. Diet + W-1372: 145 ± 10 mg/dl. |
| Arterial territories examined | Aorta, carotid bifurcations, femoral arteries, and main coronary branches. |
| Plaque-assessment method | Gross visual staining of the aortic surface, light microscopy, histomorphometry, and scanning electron microscopy. |
| Evidence of coronary disease | Advanced, occlusive fibro-lipid coronary plaques with prominent necrotic cores. |
| Evidence of plaque progression | Severe progressive plaques developed at 18 months, causing significant stenosis and media destruction. |
| Evidence of true regression | Marked reduction in gross aortic intimal lesions in the diet-only group, with further reduction when W-1372 was added. |
| Evidence of plaque stabilization | Near-complete clearance of intracellular foam cells and a reduction in necrotic debris; endothelial-surface improvements were reported, in part in companion ultrastructural studies. |
| Calcification / thrombosis / death | Intraplaque hemorrhage and focal necrosis were present at baseline and were less evident in regression animals (inferred from cross-sectional necropsy comparisons). |
| Main conclusions | Advanced atherosclerotic plaques can regress; lowering plasma cholesterol toward baseline accelerates regression and matrix stabilization, and an added pharmacologic agent augments the effect. |
| Major limitations | Quantified primarily via visual surface-area projection and light microscopy. |
| Relevance to human disease | High; supported the concept that combining lipid-lowering diet with a pharmacologic agent accelerates plaque healing and endothelial repair. |
Rank 7: Oregon Alfalfa Saponin Program (Malinow, McLaughlin, Naito, McNulty, et al., 1978)
| Full citation | [9] Malinow M. R., McLaughlin P., Naito H. K., Lewis L. A., McNulty W. P. Effect of alfalfa meal on shrinkage (regression) of atherosclerotic plaques during cholesterol feeding in monkeys. Atherosclerosis. 1978 May;30(1):27–43. |
| Year | 1978 |
| Primate species | Macaca fascicularis (Cynomolgus macaque) |
| Number of animals | 72 (a baseline-necropsy group plus three groups of ~18 during the regression phase) |
| Sex and approximate age | Adult males, age approximately 5–6 years |
| Duration of atherogenic feeding | 6 months |
| Duration of regression phase | 18 months |
| Experimental diet composition | Semipurified diet rich in saturated fat, containing sucrose, dextrin, casein, and mineral mixtures. |
| Control / regression diet | Semipurified diet with or without alfalfa meal, or a diet consisting entirely of monkey chow. |
| Dietary cholesterol content | 1.2 mg/Cal during induction; reduced during regression with or without alfalfa meal. |
| Total-fat percentage | High fat (approximately 40% of calories) during induction; low fat during chow regression. |
| Saturated-fat percentage | High saturated fat during induction. |
| Achieved lipids & lipoproteins | Induction: 734 ± 41 mg/dl (TC). Alfalfa group: 341 ± 22 mg/dl. Chow group: 146 ± 11 mg/dl. |
| Arterial territories examined | Thoracic aorta, abdominal aorta, and main coronary arteries. |
| Plaque-assessment method | Visual surface staining of the aorta and histomorphometry of cross-sectioned coronary arteries. |
| Evidence of coronary disease | Extensive proximal coronary plaques with prominent lipid-laden foam cells. |
| Evidence of plaque progression | Monkeys on the control saturated-fat diet showed progressive, severe coronary plaques over the 18-month intervention period. |
| Evidence of true regression | Alfalfa-fed monkeys showed significant reduction in aortic and coronary plaque area, approaching the chow-fed group. |
| Evidence of plaque stabilization | Plaque shrinkage was accompanied by a marked decrease in foam cell infiltration and normalization of circulating lipoproteins. |
| Calcification / thrombosis / death | No prominent treatment toxicity was reported in the publication. |
| Main conclusions | Addition of alfalfa meal was associated with lower plasma cholesterol and reduced lesion burden during continued cholesterol feeding; saponin-mediated interference with intestinal sterol absorption was proposed as a mechanism, but the intervention was a complex plant preparation rather than a purified saponin. |
| Major limitations | The active component was not isolated; a whole plant preparation was used, and the coronary change was not necessarily quantified to the same standard as the aortic change. |
| Relevance to human disease | High; supported the concept that reducing intestinal cholesterol absorption can lower plasma cholesterol and lesion burden, though the active component was not isolated. |
Rank 8: Wake Forest / Bowman Gray Fatty Acid Program (Rudel, Parks, Sawyer, 1995)
| Full citation | [10] Rudel L. L., Parks J. S., Sawyer J. K. Compared with dietary monounsaturated and saturated fat, polyunsaturated fat protects African green monkeys from coronary artery atherosclerosis. Arterioscler Thromb Vasc Biol. 1995 Dec;15(12):2101–2110. |
| Year | 1995 |
| Primate species | Chlorocebus aethiops (African green monkey; formerly Cercopithecus aethiops) |
| Number of animals | 36 |
| Sex and approximate age | Adult males, age approximately 5–7 years |
| Duration of atherogenic feeding | 5 years continuous feeding |
| Duration of regression phase | Comparative progression model (no dedicated regression phase in this cohort). |
| Experimental diet composition | Liquid-formula diet providing 35% of total kilocalories as fat, 21% from protein, and 44% from carbohydrate. |
| Control / comparison diets | Saturated fat (palm oil) vs. monounsaturated fat (oleic-enriched safflower) vs. polyunsaturated fat (standard safflower). |
| Dietary cholesterol content | 0.80 mg/kcal in all experimental formulations. |
| Total-fat percentage | 35.0% of total calories. |
| Saturated-fat percentage | Saturated group: 19.5%. Monounsaturated: 4.5%. Polyunsaturated: 5.1%. |
| Achieved lipids & lipoproteins | Saturated: 380 ± 25 (TC); Monounsaturated: 290 ± 18; Polyunsaturated: 280 ± 15 mg/dl. The monounsaturated group had the lowest LDL/HDL ratio yet did not show reduced atherosclerosis. |
| Arterial territories examined | Aorta, coronary arteries (LAD, LCx, RCA), and hepatic tissue. |
| Plaque-assessment method | Perfusion-fixed morphometry, chemical lipid-class separation, and hepatic ACAT activity assay. |
| Evidence of coronary disease | Advanced coronary atherosclerosis with prominent intimal plaque area in the saturated and monounsaturated groups. |
| Evidence of plaque progression | 5 years of palm oil or oleic acid feeding drove progressive coronary plaque development. |
| Evidence of true regression | Comparative progression study; no regression phase. Polyunsaturated fat was associated with substantially less coronary plaque than monounsaturated or saturated fat. |
| Evidence of plaque stabilization | Polyunsaturated fat minimized cholesteryl ester and cholesteryl oleate accumulation in the coronary arteries. |
| Calcification / thrombosis / death | Plaque calcification was observed in the saturated and monounsaturated groups. No acute thrombotic events were documented. |
| Main conclusions | Under this cholesterol-containing experimental diet, monounsaturated fat did not reduce coronary plaque to the degree observed with polyunsaturated fat, despite apparently favorable plasma-lipoprotein measures; polyunsaturated fat minimized cholesteryl oleate accumulation. Findings cannot be extrapolated directly to whole-food human dietary patterns. |
| Major limitations | Absence of a dedicated regression or dietary crossover phase in this cohort. |
| Relevance to human disease | Very High; challenged the assumption that monounsaturated and polyunsaturated fats have equivalent cardiovascular effects. |
Rank 9: Southwest Foundation Baboon Program (McGill, McMahan, Kruski, Mott, 1981)
| Full citation | [11] McGill H. C. Jr., McMahan C. A., Kruski A. W., Mott G. E. Relationship of lipoprotein cholesterol concentrations to experimental atherosclerosis in baboons. Arteriosclerosis. 1981 Jan–Feb;1(1):3–12. |
| Year | 1981 |
| Primate species | Papio sp. (Baboon) |
| Number of animals | 60 |
| Sex and approximate age | Young adult males and females, age approximately 4–6 years |
| Duration of atherogenic feeding | 26 months continuous feeding |
| Duration of regression phase | Baseline comparison design (diet-induced progression focus). |
| Experimental diet composition | Custom diet: 21% of calories from protein, 41% from fat, and 38% from carbohydrate. |
| Control diet | Low-fat standard baboon chow (approximately 5.8% calories from fat). |
| Dietary cholesterol content | 1.7 mg/kcal during the 26-month experimental period. |
| Total-fat percentage | 41.0% of total calories. |
| Saturated-fat percentage | 17.2% of total calories (lard and dried egg yolk). |
| Achieved lipids & lipoproteins | Chow: 112 ± 8 mg/dl (TC). Atherogenic: 298 ± 15 mg/dl (TC). |
| Arterial territories examined | Thoracic aorta, abdominal aorta, common iliac, and extramural coronary arteries. |
| Plaque-assessment method | Perfusion-fixed visual surface estimation, independent pathobiological grading, and chemical lipid extraction. |
| Evidence of coronary disease | Mild-to-moderate early-stage coronary fatty streaks and small fibrous plaques. |
| Evidence of plaque progression | 26 months of a high-saturated-fat, high-cholesterol diet drove aortic and coronary fatty-streak progression. |
| Evidence of true regression | Not directly evaluated in this study. |
| Evidence of plaque stabilization | No intervention or regression phase was tested; this was an association study. Plaque coverage was inversely correlated with HDL-cholesterol and positively with LDL+VLDL-cholesterol. |
| Calcification / thrombosis / death | No spontaneous thrombosis or myocardial infarctions reported; minimal baseline calcification observed. |
| Main conclusions | LDL + VLDL-cholesterol positively correlates, and HDL-cholesterol negatively correlates, with diet-induced plaque coverage in baboons. |
| Major limitations | Minimal development of advanced, stenotic coronary disease within 2 years. |
| Relevance to human disease | High; demonstrated associations between lipoprotein-cholesterol fractions and lesion burden under controlled conditions. These observational relationships do not establish HDL-cholesterol as a causal, modifiable treatment target. |
Rank 10: Oregon New World Program (Maruffo & Portman, 1968)
| Full citation | [2] Maruffo C. A., Portman O. W. Nutritional control of coronary artery atherosclerosis in the squirrel monkey. J Atheroscler Res. 1968 Mar–Apr;8(2):237–247. |
| Year | 1968 |
| Primate species | Saimiri sciureus (Squirrel monkey) |
| Number of animals | 24 |
| Sex and approximate age | Adult males and females, wild-caught (age unknown) |
| Duration of atherogenic feeding | 3–12 months |
| Duration of control-diet follow-up | 3–12 months |
| Experimental diet composition | Semipurified diet rich in saturated fat, containing lard and butter. |
| Control diet | Low-fat, cholesterol-free standard commercial grain-based diet. |
| Dietary cholesterol content | 0.5% to 1.0% by weight during induction; cholesterol-free during the control-diet period. |
| Total-fat percentage | High fat (approximately 35% of calories) during induction; low fat during the control period. |
| Saturated-fat percentage | High saturated fat during induction (lard and butter). |
| Achieved lipids & lipoproteins | Induction: greater than 400 mg/dl (TC); plasma cholesterol fell rapidly toward baseline after return to the control diet. |
| Arterial territories examined | Thoracic aorta, abdominal aorta, and extramural coronary arteries. |
| Plaque-assessment method | Histopathology, electron microscopy, and tissue lipid-class separation. |
| Evidence of coronary disease | Early, lipid-rich coronary atheromas with prominent intimal foam-cell accumulations. |
| Evidence of plaque progression | High-cholesterol feeding caused rapid progression of aortic and coronary fatty streaks. |
| Evidence of true regression | Return to a low-fat, cholesterol-free diet arrested progression; the extent of true anatomical regression of early lesions is characterized cautiously (see note in Methodological Critique). |
| Evidence of plaque stabilization | Reduced plaque lipid content, with thin, fibro-elastic intimal thickenings persisting. |
| Calcification / thrombosis / death | No acute coronary thrombotic events or myocardial infarctions were documented during the study. |
| Main conclusions | The study supports arrest of progression and reduced lesion lipid content after dietary normalization more clearly than it supports quantitatively proven anatomical regression of early lesions. |
| Major limitations | Small vessel caliber and physical dimension constraints limited detailed functional vascular evaluation. |
| Relevance to human disease | Moderate-High; supported the concept that early-stage coronary lesions have high metabolic plasticity. |
Dietary Interventions and Macronutrient Analysis
Analyzing the dietary interventions used in nonhuman primate studies reveals the specific nutritional drivers of atherogenesis and regression. Historical experiments systematically evaluated the interactions of total fat, saturated fat, polyunsaturated fat, and cholesterol, providing insights that go beyond simple “high-fat” versus “low-fat” categorization.
Saturated vs. Polyunsaturated Fatty Acids in Progression and Regression
A primary question of the 1970s and 1980s was whether reversing atherosclerosis required a very-low-fat diet, or if a diet rich in polyunsaturated fat could achieve similar outcomes. In the landmark rhesus macaque study by Armstrong et al., the progression phase was driven by a diet containing 41% of total calories from fat, primarily from dried egg yolk, which produced severe, diffuse coronary stenosis.
During the 40-month regression phase, investigators compared a low-fat diet (4% of total calories from fat, primarily from grain starch) with a high-polyunsaturated-fat diet (40% of total calories from corn oil, rich in linoleic acid). Both diets were entirely cholesterol-free. The outcomes of this head-to-head comparison were striking:
- Both diets reduced plasma total cholesterol to nearly identical levels (approximately 140 mg/dl).
- Both diets reversed the atherogenic beta-lipoprotein-dominant (LDL) profile back toward alpha-lipoprotein dominance (the historical electrophoretic correlate of the HDL fraction, which is not identical to today’s HDL subclasses).
- Anatomical coronary regression was similar between the groups: the average cross-sectional lumen area was more than 80% greater in regression animals than in animals with baseline disease, with no significant difference between the low-fat and corn-oil diets. The decisive shared feature was not the total-fat content but the fact that both diets normalized circulating cholesterol, LDL, and (by inference) the ApoB-particle burden; regression followed that normalization.
In this model, very low total fat was not necessary for regression: a cholesterol-free, polyunsaturated-fat-rich diet produced an equivalent and sustained reduction in circulating atherogenic lipoproteins and comparable regression. The experiment therefore identifies the achieved lipoprotein response—rather than total-fat percentage alone—as the factor most closely associated with regression. It does not establish that total dietary fat is irrelevant across other species, fatty-acid compositions, energy intakes, or ordinary human diets; the effects of saturated fat and dietary cholesterol on circulating ApoB are substantial but not deterministic.
The Confounding Roles of Casein, Sucrose, and Caloric Intake
Evaluating historical primate experiments requires separating the effects of dietary fat and cholesterol from other ingredients in semipurified diets. Many atherogenic formulas contained casein as the sole protein source and sucrose as the primary carbohydrate, both of which can have independent metabolic effects.
Casein has been reported to raise plasma cholesterol in several mammalian species compared with soy or other plant proteins, an effect that varies considerably by species and is possibly related to differences in amino-acid composition and in cholesterol and bile-acid metabolism. Refined sucrose does not by itself produce atherosclerotic plaque, but high intakes raise hepatic VLDL synthesis, plasma triglycerides, and remnant lipoproteins and may worsen an atherogenic lipoprotein profile. Furthermore, many primate studies allowed ad libitum feeding, resulting in positive energy balance and weight gain, which can confound metabolic assessments. A related caution applies to the cholesterol-free regression diets used across these programs: because such diets often changed fat source, fat percentage, energy density, protein source, fiber, and plant compounds simultaneously, readers should not infer that dietary cholesterol alone accounts for either disease induction or its reversal.
However, in trials that controlled for these variables—such as the LSU rhesus program, which used identical casein, sucrose, and fat levels in both the progression and regression feeds, varying only the cholesterol content—deleting cholesterol while holding saturated fat high was sufficient to lower plasma cholesterol and induce regression. This underscores the central role of dietary cholesterol in this model, while not excluding independent contributions of saturated fat that have been demonstrated in other experimental designs.
Progression and Regression Dynamics: Plaque vs. Stabilization
Diet-induced atherosclerosis in nonhuman primates follows a predictable pathological sequence that closely mirrors the development of human coronary artery disease.
ApoB-particle entry & retention → aggregation & biochemical modification → endothelial / innate immune activation → lipid loading of macrophage- and smooth-muscle-derived foam cells → extracellular lipid, matrix remodeling, cell death & defective efferocytosis → fibrous cap / necrotic core
Pathological Milestones of Progression
- Endothelial Activation & Fatty Streaks: Elevated circulating ApoB particles are retained in the subendothelial space, where they undergo aggregation, oxidation, and other biochemical modifications. This triggers endothelial adhesion molecules and recruits blood-derived monocytes, which differentiate into macrophages and internalize modified lipoproteins to become foam cells, forming early fatty streaks.
- Fibrous Plaque Development: Under continued hypercholesterolemia, foam cells release growth factors (such as platelet-derived growth factor) that stimulate smooth muscle cell migration from the media. These cells proliferate and secrete extracellular matrix proteins, creating a fibrous cap over the lipid-rich core; smooth-muscle-derived cells can themselves take up lipid and contribute substantially to the foam-cell population.
- Necrotic Core Formation: As the plaque matures, foam cells within the core undergo apoptosis and secondary necrosis. Failure of efferocytosis leads to accumulation of cellular debris, free cholesterol, and lipid pools, forming a highly unstable necrotic core.
- Calcification and Media Changes: Microscopic calcium granules emerge within the necrotic core and degenerating smooth muscle cells and coalesce into large, rigid mineral plates. The adjacent tunica media often exhibits cellular infiltration, elastic lamina fragmentation, and focal thinning.
Plaque Stabilization vs. Anatomical Shrinkage
- Macrophage Depletion and Inflammatory Resolution: Marked lipid lowering reduces continued monocyte recruitment and promotes the loss, clearance, and phenotypic remodeling of plaque macrophages; the relative contributions of emigration, cell death, and efferocytosis vary by model and lesion stage. Macrophages downregulate inflammatory genes and upregulate tissue-remodeling markers, and necrotic debris is progressively removed as local inflammation resolves.
- Matrix Remodeling & Fiber Cross-Linking: While lipid and cellular components are cleared comparatively more quickly, extracellular matrix elements (collagen, elastin) are remodeled more slowly. Smooth muscle cells synthesize new collagen fibers that cross-link and condense, transforming a soft, unstable atheroma into a flatter, more stable, fibrous lesion.
- Endothelial Healing: The endothelial lining over remaining plaque recovers structural integrity, forming continuous, tightly joined cell junctions that resist shear stress and reduce thrombogenicity.
These changes indicate that plaque stabilization and structural healing precede, and can occur independently of, a significant reduction in overall plaque size.
In modern mechanistic terms, the removal of plaque lipid during regression is understood to depend heavily on reverse cholesterol transport: efflux of free cholesterol from lipid-laden cells via the transporters ABCA1 and ABCG1 to HDL acceptors, esterification by lecithin–cholesterol acyltransferase (LCAT), and ultimate hepatic clearance. These efflux pathways were not directly measured in the historical primate experiments, but they provide the contemporary biochemical framework for the lipid depletion those studies documented morphologically and chemically.
Macromolecular and Kinetic Mechanisms (ApoB and the Cumulative-Exposure Model)
ApoB-Containing Lipoproteins and Cumulative Arterial Exposure
The entry and subsequent retention of ApoB-containing lipoproteins within the subendothelial space constitute the fundamental initiating lipid event in atherosclerotic plaque formation. LDL, intermediate-density lipoproteins, cholesterol-enriched remnants, and lipoprotein(a) can cross the arterial endothelium. Once within the intima, positively charged regions of apolipoprotein B interact with negatively charged glycosaminoglycan chains on arterial-wall proteoglycans (notably biglycan and versican), selectively retaining these particles at lesion-prone sites [12]. Retained particles undergo aggregation, enzymatic modification, oxidation, and other compositional changes that provoke innate and adaptive immune responses, macrophage foam-cell formation, smooth-muscle-cell phenotypic change, extracellular lipid accumulation, and ultimately formation of a fibrous and potentially necrotic plaque. Because each atherogenic LDL, IDL, remnant, or Lp(a) particle carries a single ApoB molecule, it is the number of retained particles—not the cholesterol mass they carry—that is mechanistically trapped in the wall; this is why particle-based (ApoB) measures are conceptually preferable to cholesterol-mass (LDL-C) measures, even though the historical studies could report only the latter.
Hemodynamic forces, endothelial transcytosis, arterial proteoglycan composition, inflammation, and genetic susceptibility strongly influence the location and rate of lesion development. These factors do not displace ApoB-containing particles from the causal center of atherogenesis; rather, they regulate the probability that circulating particles will enter, remain within, and injure a particular arterial region. In this framework, ApoB particle burden supplies the atherogenic substrate, arterial-wall characteristics govern retention and biological response, and inflammatory and thrombotic pathways determine plaque progression and clinical complication.
The progression of atherosclerosis is therefore influenced by cumulative arterial exposure to circulating ApoB-containing particles. This relationship may be represented conceptually as:
Cumulative Exposure = ∫0t CApoB(t) dt
where C_ApoB(t) is the circulating concentration of atherogenic ApoB-containing particles over time. The equation is not intended to imply that identical calculated exposure produces identical plaque in every individual. The biological consequences of a given exposure are modified by particle composition, arterial-wall retention affinity, age, blood pressure, smoking, glycemia, inflammation, hemodynamics, and genetic susceptibility. Nevertheless, the primate literature is consistent with, and provides experimental support for, the cumulative ApoB-exposure model derived from modern human genetic, epidemiologic, and clinical evidence [13], [14]: longer and greater exposure produces older, more fibrotic, calcified, and metabolically resistant lesions, whereas early lipid-rich lesions remain substantially more reversible.
Most historical primate studies measured plasma total cholesterol, beta-lipoprotein cholesterol, or LDL-plus-VLDL cholesterol rather than ApoB directly. Their findings can nevertheless be interpreted within the modern ApoB framework because the atherogenic diets produced marked increases in circulating ApoB-containing lipoproteins, while successful regression regimens produced large and sustained reductions in those particles. The close relationship between normalization of the atherogenic lipoprotein burden and plaque arrest, lipid depletion, stabilization, and partial anatomical regression provides experimental support for the causal importance of cumulative ApoB exposure.
The LSU long-term rhesus studies illustrate this principle: animals exposed to a sustained lipid challenge over 5.4 years developed highly complex, fibrotic, structured plaques that needed a substantially longer duration of lipid lowering (regression becoming demonstrable at approximately 3.7 years) than early lipid-rich lesions, indicating that the physical age of a plaque, combined with its cumulative exposure history, influences its metabolic reversibility.
Comparative Analysis with Human Pathology and Clinical Trials
Translating the pathological findings of historical primate experiments to human clinical practice requires a careful evaluation of the shared pathways and structural differences between these models and human disease.
Structural Parallels and Shared Mechanisms
The cellular and molecular mechanisms of atherogenesis are highly conserved between humans and nonhuman primates. Both species initiate plaque formation through the subendothelial retention and oxidation of ApoB-containing lipoproteins. Subsequent steps—including monocyte migration, differentiation into macrophages, scavenger-receptor-mediated lipid uptake, smooth muscle cell migration, and necrotic core formation—are closely similar in both species. Under prolonged dietary challenge, both rhesus and cynomolgus macaques develop advanced plaques characterized by fibrous caps, lipid cores, calcification, and media degradation, resembling advanced (complicated) human atherosclerotic lesions. Although histologically similar, experimentally induced monkey plaques typically remain simpler than many decades-old human plaques that have been exposed to multiple, concurrent cardiovascular risk factors.
Shared Clinical and Pathological Consequences
- Myocardial Infarction: Rare spontaneous, fatal myocardial infarctions were reported in some long-term, severely hypercholesterolemic macaque colonies. A frequently cited estimate is roughly 1 per 300 monkeys at risk per year [15], [16]; this figure derives from a commentary rather than a directly analyzed incidence cohort and should not be generalized across species or protocols. Where they occurred, these infarctions were morphologically similar to human infarcts, showing transmural or subendocardial necrosis, contraction band necrosis, and associated electrocardiographic changes.
- Vascular Reactivity & Vasomotor Tone: Both species exhibit impaired endothelial nitric oxide (NO) bioavailability during atherogenesis. Under hypercholesterolemic conditions, coronary arteries lose their ability to dilate in response to acetylcholine (instead exhibiting paradoxical vasoconstriction), a defect that, in studies that directly measured vascular reactivity, was reversible upon sustained lipid lowering.
- Cerebrovascular & Peripheral Disease: Monkeys fed atherogenic diets develop atherosclerosis in extracoronary arteries such as the carotid bifurcation and the femoral arteries, mimicking the systemic distribution of human arterial disease.
Plaque Remodeling: Primate vs. Human Imaging Outcomes
Evaluating regression in human clinical trials relies on non-invasive imaging technologies that assess specific dimensions of plaque biology. Nonhuman primate studies provide the histopathological verification that underpins the interpretation of these clinical endpoints.
- Lumen Diameter (QCA): Early human trials used quantitative coronary angiography to measure change in luminal diameter, occasionally documenting small improvements. Quantitative coronary angiography records luminal dimensions and cannot by itself distinguish plaque shrinkage from changes in arterial remodeling (the outward, plaque-compensating enlargement first characterized by Glagov [17]) or vasomotor tone; primate studies show that lipid lowering can improve lumen dimensions and vascular reactivity without a proportionate reduction in histological plaque area.
- Total Plaque Volume (IVUS & OCT): Modern trials use intravascular ultrasound and optical coherence tomography to measure changes in total plaque volume, often showing small reductions. Primate studies suggest these reductions represent cellular and chemical clearance of lipid pools and necrotic debris from the plaque core.
- Noncalcified Plaque Changes (CCTA): Coronary CT angiography allows non-invasive quantification of soft, noncalcified plaque volume. Primate data indicate this pool contains highly active, lipid-laden foam cells and extracellular lipid deposits, which are the most rapidly cleared components during regression. Consistent with modern IVUS and CCTA observations, an increase in calcium density can accompany this loss of lipid-rich plaque during healing, so a rising calcium signal does not necessarily indicate disease progression.
- Vulnerable Plaque Features & Healing: OCT and PET imaging in humans track fibrous-cap thickening, necrotic-core reduction, and stabilization of inflamed plaques. Nonhuman primate histopathology is consistent with these findings, showing improved endothelial coverage, reduced foam-cell populations, and deposition of a denser, more stable collagen cap over remaining necrotic debris; whether foam-cell loss reflects emigration, local cell death, reduced recruitment, or efflux was not resolved in most historical studies.
The primate findings sit within a broader modern framework. The response-to-retention hypothesis formalized subendothelial ApoB-particle retention as the initiating step [12]; human Mendelian-randomization and epidemiologic work established cumulative LDL/ApoB exposure as a determinant of lifetime risk [13], [14]; and the inflammatory contribution to events was confirmed clinically in the CANTOS trial [18]. In parallel, serial intravascular-imaging and CT trials of intensive lipid lowering—REVERSAL [19], ASTEROID [20], SATURN [21], GLAGOV [22], PACMAN-AMI [23], and HUYGENS [24]—have documented plaque regression, stabilization, and compositional change in living patients, connecting the historical primate evidence to contemporary human data.
Methodological and Ethical Critique
Evaluating historical primate literature requires a critical analysis of both the experimental designs of the mid-to-late twentieth century and the evolution of contemporary animal welfare regulations.
Methodological Limitations and Design Bias
- Cross-Sectional Postmortem Comparisons: Most historical regression studies relied on a cross-sectional design in which a subgroup of animals was necropsied at the end of induction to establish “baseline progression,” and the remaining animals were randomized to regression arms and evaluated years later. This design assumes the baseline group is representative of the surviving regression animals. Given the high individual variation in dietary response (the “hyper-responder” versus “hypo-responder” phenomenon), it can introduce significant statistical noise and bias.
- Survivorship Bias: In long-term progression studies, animals with the most severe coronary stenosis were at high risk for sudden cardiac death before the formal regression phase, which can lead to selective evaluation of more resistant, hypo-responsive animals in the final regression cohorts.
- Small Sample Sizes: Because of the high cost and complexity of primate husbandry, many cohorts were small (often 6 to 12 per arm), limiting statistical power and complicating subgroup analyses.
- Colony Overlap and Duplicate Reporting: Investigators frequently generated multiple, overlapping publications from the same cohorts over several years, making it difficult to isolate independent experimental results in systematic reviews.
- Note on Maruffo & Portman (1968): Secondary summaries of this early squirrel-monkey work describe a comparatively short atherogenic period followed by a control-diet period in which lesions did not progress but did not clearly regress. The extent of true anatomical regression in this specific study should therefore be characterized cautiously and confirmed against the primary report.
Historical Terminology vs. Modern Usage
- “Cholesterol Atherosclerosis” & “Atheromatosis”: Historically used to describe the diffuse lipid-loading and foam cell infiltration driven by extreme dietary cholesterol feeding. Modern pathology reserves “atherosclerosis” for the complex, chronic, inflammatory-fibromuscular disease process, distinguishing it from simple, non-inflammatory intimal lipid deposition.
- “Beta-Lipoproteinemia”: Refers to the historical electrophoretic classification of circulating lipoproteins, where beta-lipoproteins corresponded to the LDL fraction and alpha-lipoproteins to the HDL fraction. These electrophoretic bands are not identical to the density-defined subclasses of modern lipidology; contemporary biochemistry uses density ultracentrifugation and immunoassays to directly quantify specific apolipoprotein particles, particularly ApoB and ApoA-I. The historical primate targets (for example, a total-cholesterol ceiling near 200 mg/dl) also predated routine ApoB measurement.
- “Regression”: Historically defined as any reduction in visual arterial lesion coverage or tissue cholesterol content. Modern vascular biology distinguishes true volumetric regression from structural stabilization, fibrous-cap reinforcement, and functional vasomotor recovery.
Ethical Evolution of Primate Research Regulations
The regulatory landscape governing animal research underwent a profound transformation during this era. The original United States Laboratory Animal Welfare Act of 1966 was primarily enacted to regulate the transport, sale, and handling of six species, including nonhuman primates, to prevent pet theft and ensure basic humane care. A 1970 amendment renamed the statute the Animal Welfare Act and broadened coverage toward warm-blooded animals used in research, with a further amendment in 1976.
The most extensive regulatory shift occurred with the 1985 Improved Standards for Laboratory Animals Act, enacted as part of the Food Security Act of 1985. This amendment introduced requirements that altered the design of primate research:
- Institutional Animal Care & Use Committees (IACUCs): Mandated independent review boards to evaluate and approve experimental protocols involving covered species, ensuring that pain and distress are minimized.
- Environmental Enrichment & Psychological Well-Being: Required institutions to develop plans promoting the psychological well-being of nonhuman primates, with environmental enrichment and, where appropriate, compatible social housing among the preferred measures rather than an unconditional requirement for every animal.
- Reduction & Alternatives: Compelled researchers to formally consider alternatives to painful procedures and to minimize the number of animals used.
Simultaneously, the Health Research Extension Act of 1985 amended the Public Health Service Act and provided the statutory basis for the PHS Policy on Humane Care and Use of Laboratory Animals, enforcing compliance with the Guide for the Care and Use of Laboratory Animals for NIH-funded institutions. These regulatory changes, combined with rising financial costs and international conservation measures, sharply curtailed the large-scale, invasive primate experiments of the mid-twentieth century. Modern research relies heavily on non-invasive imaging and highly refined, ethically approved protocols.
Explicit Investigation of Core Hypotheses and Counter-Evidence
To evaluate the scientific integrity of the primate literature, the core hypotheses of this field are examined below alongside the evidence that supports and challenges each.
Hypothesis 1: Nonhuman primates develop coronary atherosclerosis that is histologically and metabolically closer to human disease than most other laboratory animals.
Support. Rhesus and cynomolgus macaques develop complex epicardial coronary plaques featuring fibrous caps, lipid-rich necrotic cores, calcification, media degradation, and spontaneous myocardial infarctions that closely match human lesions.
Counter-Evidence / Challenges. Certain species, such as baboons and squirrel monkeys, are relatively resistant to severe, occlusive coronary stenosis or advanced plaque rupture within standard timeframes, limiting their utility for modeling late-stage clinical events.
Hypothesis 2: Diet-induced elevations in ApoB-containing lipoproteins are the principal mediator between atherogenic diets and plaque formation.
Support. Plaque progression and regression correlate strongly with circulating VLDL+LDL cholesterol, and clearing these ApoB-containing particles is required to arrest and reverse plaque development.
Counter-Evidence / Challenges. Local proteoglycan binding, hemodynamics, and chemical modifications (such as oxidation) are also required to initiate plaque formation. Some animals show significant variation in lesion severity at identical circulating ApoB levels, indicating individual genetic and vascular susceptibility.
Hypothesis 3: Early lipid-rich lesions can regress substantially after major cholesterol reduction.
Support. Early fatty streaks and foam-cell-rich lesions in rhesus and squirrel monkeys undergo rapid and substantial regression within months following a return to low-cholesterol diets.
Counter-Evidence / Challenges. In advanced, highly structured lesions, simple lipid lowering may clear intracellular cholesteryl esters but does not rapidly reduce overall plaque cross-sectional area, as the mature collagenous and calcified matrix remains largely unchanged.
Hypothesis 4: Advanced fibrotic or calcified lesions regress incompletely but may become more stable.
Support. Long-term regression studies show that while advanced, years-old plaques lose inflammatory cell content and soft lipid pools, dense collagenous scars and large calcified plates persist within the arterial wall.
Counter-Evidence / Challenges. Under certain long-term protocols, very slow reductions in plaque calcium and enzymatic degradation of collagen have been reported, indicating that even mature matrix components possess slow, long-term turnover.
Hypothesis 5: The duration and cumulative magnitude of lipoprotein exposure influence whether plaque is reversible.
Support. LSU studies showed that regression of plaques induced over 5.4 years became statistically demonstrable only after an extended regression phase (approximately 3.7 years), in contrast to the more rapid response of early fatty streaks.
Counter-Evidence / Challenges. In some experiments, genetic “hypo-responder” animals showed significant regression under moderate lipid lowering, whereas “hyper-responders” progressed, indicating that individual metabolic susceptibility can override cumulative-exposure targets.
Hypothesis 6: Very-low-fat diets promote regression primarily when they produce large and sustained reductions in atherogenic lipoproteins.
Support. Reversing hypercholesterolemia using ultra-low-fat diets (4% of calories as fat) lowers plasma cholesterol and regresses coronary plaques.
Counter-Evidence / Challenges. Corn-oil-substitution diets (40% of calories as fat) achieve comparable plasma cholesterol lowering and coronary regression when dietary cholesterol is eliminated, indicating that absolute lipid clearance, rather than total dietary fat reduction, is the primary driver.
Hypothesis 7: Human coronary regression is generally smaller and slower than in some monkey studies because human plaques are older, more heterogeneous, and exposed to multiple risk factors.
Support. Human plaques accumulate over decades and are characterized by highly cross-linked, mature collagen and calcification. Diet-induced monkey lesions are typically generated over 1 to 5 years and contain younger, more metabolically active lipids that are more easily cleared.
Counter-Evidence / Challenges. Aggressive modern pharmacological regimens can lower human LDL-cholesterol to very low levels, which has been associated with rapid regression of soft, noncalcified lipid cores, approaching the kinetics seen in some primate studies.
Hypothesis 8: Clinical benefit may result more from plaque stabilization and prevention of new lesions than from complete removal of existing plaque.
Support. Primate lipid-lowering trials demonstrated an approximate doubling of coronary lumen size via outward remodeling and improved endothelium-dependent vasomotor dilation to acetylcholine, despite persistence of the advanced calcified plaque matrix.
Counter-Evidence / Challenges. While stabilization reduces acute ischemic events, more complete anatomical reduction of plaque may still be needed to restore normal mechanical vascular compliance and relieve severe, chronic exertional angina in highly stenotic vessels.
What These Animal Studies Do Not Prove
To ensure rigorous translation to human clinical practice, the scientific and biological limits of nonhuman primate trials must be clearly defined:
- They do not prove that diet alone can reverse advanced coronary artery disease in all humans. Primate studies used highly controlled, extreme dietary changes (such as shifting from a high-fat, high-cholesterol diet to an entirely cholesterol-free diet) under laboratory conditions that are difficult for human patients to achieve or sustain.
- They do not prove that coronary artery calcium will disappear. Historical pathology demonstrated that while lipid-rich pools and cell debris can be cleared, large, consolidated calcific deposits persist within the vessel wall, undergoing little to no anatomical regression over several years of lipid normalization. Calcification is not a single quantity: frequency, calcified area, microcalcification, macrocalcified plates, and calcium density can change independently, and in human therapy an increase in calcium density can accompany reduction of lipid-rich plaque and lower event rates, so persistent or increasing calcium does not by itself indicate treatment failure.
- They do not prove that human plaques regress at the same speed as monkey plaques. Nonhuman primates have significantly shorter lifespans and higher metabolic rates. A 3-year regression period in a rhesus monkey represents a substantial fraction of its lifespan, whereas human plaques develop over decades and are exposed to multiple, often unmanaged risk factors.
- They do not prove the human safety or efficacy of high-dose alternative agents. While plant-derived saponins or alfalfa meal induced regression in cynomolgus monkeys by blocking intestinal absorption, such agents can cause systemic toxicities (such as hemolytic effects) in humans if not carefully purified, and they are not approved for clinical use.
Unanswered Questions and Contemporary Experimental Opportunities
Critical Unanswered Questions
- What is the precise phenotypic fate of plaque smooth muscle cells during dietary regression, and do they revert from a synthetic, proliferative state back to a contractile state?
- How does systemic inflammation (measured by high-sensitivity C-reactive protein or key interleukins) interact with aggressive lipid-lowering diets to determine the speed of primate coronary regression?
- Can advanced coronary microvascular dysfunction be fully reversed by intensive dietary modification alone, or does it require concurrent pharmacotherapy?
Contemporary, Highly Ethical Experimental Designs
Studies involving prolonged severe hypercholesterolemia, substantial morbidity, or terminal pathological assessment would now face far more demanding scientific-necessity, harm–benefit, refinement, veterinary-monitoring, and IACUC review; such work is not categorically prohibited, but many historical protocols would be difficult to justify or would require substantial redesign. Within these constraints, highly informative, minimally invasive studies can be conducted in nonhuman primates today:
- Serial, Non-Invasive High-Resolution Imaging: High-resolution MRI, PET/CT, and coronary CT angiography can track progression and regression within the same living animals over time, eliminating the statistical noise of cross-sectional designs and enabling real-time evaluation of plaque volume, lipid content, and calcification.
- Pedigreed, Aged Nonhuman Primate Colonies: Established, naturally occurring hyperlipidemic or aged primate cohorts allow evaluation of gene-diet interactions and the impact of aging on plaque stability without invasive surgery or rapid, unphysiological lipid-loading regimens.
- Peripheral Blood Transcriptomics and Biomarkers: Single-cell RNA sequencing of peripheral blood mononuclear cells and circulating inflammatory markers during dietary interventions can map the systemic immune and metabolic pathways that drive vascular healing and reverse cholesterol transport.
Central Synthesis and Answer to the Central Question
Question. To what extent do historical monkey experiments demonstrate that human-like coronary atherosclerosis caused by prolonged exposure to elevated atherogenic lipoproteins can be slowed, stabilized, or partially reversed through major dietary and plasma-lipid reduction, and what aspects of those findings can and cannot reasonably be extrapolated to human coronary artery disease?
Historical nonhuman primate experiments between 1950 and the late 1980s provide histologically verified evidence that human-like coronary atherosclerosis is a dynamic and potentially reversible pathology. These trials demonstrate that when severe hypercholesterolemia is aggressively reversed, the progression of coronary atheromas is arrested and substantial anatomical regression can be achieved. Early, lipid-rich plaques possess high metabolic plasticity, allowing rapid clearance of intracellular foam cells, hydrolysis of cholesteryl esters, and resolution of early-stage stenosis within months.
By contrast, advanced, calcified, and densely fibrotic plaques regress much more slowly and incompletely, leaving a persistent collagenous scar and mineral deposits within the vessel wall. Crucially, these studies show that the clinical benefits of lipid lowering are not restricted to physical plaque shrinkage. Substantial benefit results from plaque stabilization—including lipid depletion, reduced cellularity and inflammation, increased relative collagen content, endothelial healing, and (in modern human imaging studies) fibrous-cap thickening—together with enlargement of coronary artery and lumen cross-sectional area consistent with favorable remodeling and restored endothelium-dependent vasomotor dilation.
Extrapolatable Findings
- The Shared Cellular Mechanics of Reversal: The core pathways of ApoB retention, foam-cell reduction, cholesterol efflux, fibrous-cap reinforcement, and endothelial healing are broadly conserved between primates and humans, supporting the biological plausibility of clinical plaque stabilization.
- The Targets of Volumetric Change: Soft, noncalcified, lipid-rich plaque cores are the most rapidly and completely cleared components during regression, matching human intravascular imaging findings.
- Outward Remodeling & Functional Vasomotor Recovery: The primate finding that lipid lowering can approximately double the coronary lumen area and restore endothelial reactivity helps explain why human cardiovascular event rates decline following lipid-lowering therapy despite minimal overall change in angiographic stenosis.
Non-Extrapolatable Findings
- The Kinetics and Speed of Regression: Human coronary plaques are older, more structurally complex, and characterized by highly cross-linked, mature collagen and calcification. They are less metabolically active and regress much more slowly than the younger, diet-induced lesions of experimental primates.
- The Complexity of Multi-Risk Etiologies: Primate trials examined a single, severe, diet-induced risk factor under uniform conditions. Human coronary artery disease is multifactorial, driven by the interaction of lipid levels with cigarette smoking, hypertension, diabetes, and genomic variation.
- The Feasibility of Extreme Dietary Reversals: The extreme dietary changes used to drive regression in primate trials are difficult to achieve or sustain in clinical practice, requiring human therapies to rely on a combination of moderate dietary modification and intensive pharmacological lipid-lowering.
- No Endorsement of a Specific Named Diet: These experiments do not establish that any particular human dietary program—Ornish, Esselstyn, vegan, or Mediterranean—is superior. What they demonstrate is narrower and mechanism-based: aggressive, sustained reduction of atherogenic (ApoB-containing) lipoproteins, by whatever means, can arrest progression and induce regression under controlled conditions.
References
- TAYLOR CB, PATTON DE, COX GE. ATHEROSCLEROSIS IN RHESUS MONKEYS. VI. FATAL MYOCARDIAL INFARCTION IN A MONKEY FED FAT AND CHOLESTEROL. Arch Pathol. 1963;76:404-412.
- Maruffo CA, Portman OW. Nutritional control of coronary artery atherosclerosis in the squirrel monkey. J Atheroscler Res. 1968;8(2):237-247. doi:10.1016/s0368-1319(68)80060-2
- Armstrong ML, Warner ED, Connor WE. Regression of coronary atheromatosis in rhesus monkeys. Circ Res. 1970;27(1):59-67. doi:10.1161/01.res.27.1.59
- Weber G, Fabbrini P, Resi L, Jones R, Vesselinovitch D, Wissler RW. Regression of arteriosclerotic lesions in rhesus monkey aortas after regression diet. Scanning and transmission electron microscope observations of the endothelium. Atherosclerosis. 1977;26(4):535-547. doi:10.1016/0021-9150(77)90121-6
- Clarkson TB, Bond MG, Bullock BC, McLaughlin KJ, Sawyer JK. A study of atherosclerosis regression in Macaca mulatta. V. Changes in abdominal aorta and carotid and coronary arteries from animals with atherosclerosis induced for 38 months and then regressed for 24 or 48 months at plasma cholesterol concentrations of 300 or 200 mg/dl. Exp Mol Pathol. 1984;41(1):96-118. doi:10.1016/0014-4800(84)90011-x
- Small DM, Bond MG, Waugh D, Prack M, Sawyer JK. Physicochemical and histological changes in the arterial wall of nonhuman primates during progression and regression of atherosclerosis. J Clin Invest. 1984;73(6):1590-1605. doi:10.1172/JCI111366
- Williams JK, Anthony MS, Honoré EK, et al. Regression of atherosclerosis in female monkeys. Arterioscler Thromb Vasc Biol. 1995;15(7):827-836. doi:10.1161/01.atv.15.7.827
- Strong JP, Bhattacharyya AK, Eggen DA, Malcom GT, Newman WP 3rd, Restrepo C. Long-term induction and regression of diet-induced atherosclerotic lesions in rhesus monkeys. I. Morphological and chemical evidence for regression of lesions in the aorta and carotid and peripheral arteries. Arterioscler Thromb. 1994;14(6):958-965. doi:10.1161/01.atv.14.6.958
- Malinow MR, McLaughlin P, Naito HK, Lewis LA, McNulty WP. Effect of alfalfa meal on shrinkage (regression) of atherosclerotic plaques during cholesterol feeding in monkeys. Atherosclerosis. 1978;30(1):27-43. doi:10.1016/0021-9150(78)90150-8
- Rudel LL, Parks JS, Sawyer JK. Compared with dietary monounsaturated and saturated fat, polyunsaturated fat protects African green monkeys from coronary artery atherosclerosis. Arterioscler Thromb Vasc Biol. 1995;15(12):2101-2110. doi:10.1161/01.atv.15.12.2101
- McGill HC Jr, McMahan CA, Kruski AW, Mott GE. Relationship of lipoprotein cholesterol concentrations to experimental atherosclerosis in baboons. Arteriosclerosis. 1981;1(1):3-12. doi:10.1161/01.atv.1.1.3
- Williams KJ, Tabas I. The response-to-retention hypothesis of early atherogenesis. Arterioscler Thromb Vasc Biol. 1995;15(5):551-561. doi:10.1161/01.atv.15.5.551
- Ference BA, Ginsberg HN, Graham I, et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur Heart J. 2017;38(32):2459-2472. doi:10.1093/eurheartj/ehx144
- Borén J, Chapman MJ, Krauss RM, et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: pathophysiological, genetic, and therapeutic insights: a consensus statement from the European Atherosclerosis Society Consensus Panel. Eur Heart J. 2020;41(24):2313-2330. doi:10.1093/eurheartj/ehz962
- Clarkson TB, Mehaffey MH. Coronary heart disease of females: lessons learned from nonhuman primates. Am J Primatol. 2009;71(9):785-793. doi:10.1002/ajp.20693
- Bond MG, Bullock BC, Bellinger DA, Hamm TE. Myocardial infarction in a large colony of nonhuman primates with coronary artery atherosclerosis. Am J Pathol. 1980;101(3):675-692.
- Glagov S, Weisenberg E, Zarins CK, Stankunavicius R, Kolettis GJ. Compensatory enlargement of human atherosclerotic coronary arteries. N Engl J Med. 1987;316(22):1371-1375. doi:10.1056/NEJM198705283162204
- Ridker PM, Everett BM, Thuren T, et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N Engl J Med. 2017;377(12):1119-1131. doi:10.1056/NEJMoa1707914
- Nissen SE, Tuzcu EM, Schoenhagen P, et al. Effect of intensive compared with moderate lipid-lowering therapy on progression of coronary atherosclerosis: a randomized controlled trial. JAMA. 2004;291(9):1071-1080. doi:10.1001/jama.291.9.1071
- Nissen SE, Nicholls SJ, Sipahi I, et al. Effect of very high-intensity statin therapy on regression of coronary atherosclerosis: the ASTEROID trial. JAMA. 2006;295(13):1556-1565. doi:10.1001/jama.295.13.jpc60002
- Nicholls SJ, Ballantyne CM, Barter PJ, et al. Effect of two intensive statin regimens on progression of coronary disease. N Engl J Med. 2011;365(22):2078-2087. doi:10.1056/NEJMoa1110874
- Nicholls SJ, Puri R, Anderson T, et al. Effect of Evolocumab on Progression of Coronary Disease in Statin-Treated Patients: The GLAGOV Randomized Clinical Trial. JAMA. 2016;316(22):2373-2384. doi:10.1001/jama.2016.16951
- Räber L, Ueki Y, Otsuka T, et al. Effect of Alirocumab Added to High-Intensity Statin Therapy on Coronary Atherosclerosis in Patients With Acute Myocardial Infarction: The PACMAN-AMI Randomized Clinical Trial. JAMA. 2022;327(18):1771-1781. doi:10.1001/jama.2022.5218
- Nicholls SJ, Kataoka Y, Nissen SE, et al. Effect of Evolocumab on Coronary Plaque Phenotype and Burden in Statin-Treated Patients Following Myocardial Infarction. JACC Cardiovasc Imaging. 2022;15(7):1308-1321. doi:10.1016/j.jcmg.2022.03.002